Goal: We seek to understand
the circuitry of the mammalian cerebral cortex and how it endows us with
the ability to see.
Approach: We study the visual
cortex of both monkeys and rodents. With monkeys, we take advantage of
their ability to perform difficult perceptual tasks that provide insight
into specific aspects of their visual experience. This allows us to define
the neural correlates of particular visual behaviors and then begin to
study their underlying mechanisms, first at a coarse level, by activating or
inactivating higher order visual areas. In rodents, we exploit the ability
to continuously monitor neural populations while animals experience
“binocular rivalry” and other interesting perceptual states. We then use optogenetics to manipulate specific cell types, in
order to test hypotheses generated by the behavioral experiments in both
species.
Techniques: In monkeys, our primary tools are extracellular
electrophysiology—with multi-electrode arrays and laminar probes—and
psychophysics. They are complemented by techniques that allow us to perturb
visual areas, including inactivation with cortical cooling and activation
with microstimulation. In rodents we use
behavior, 2-photon Ca++ imaging and optogenetics.
|
|
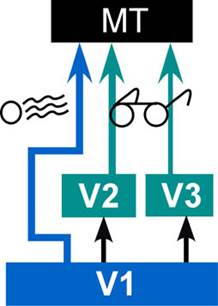
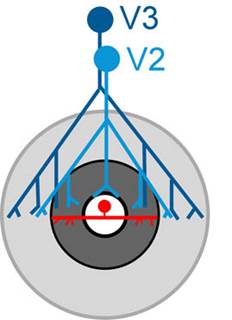
Cortico-cortical feedback. Most
of the projects in the lab are aimed at deciphering this ubiquitous, but
poorly understood, aspect of cortical connectivity. We have previously
shown that feedback from V2 and V3 has a relatively selective effect on the
non-classical surrounds of V1 receptive fields (Nassi et al. 2013;
Nassi et al. 2014). These surrounds are critical
for vision, because they allow local, feature-selective responses to be
modulated by the context in which they occur. This modulation is
surprisingly sophisticated, and appears well suited to reduce redundancy
and create sparse representations in visual cortex via input-gain control (Trott & Born 2015).
In addition, feedback exerts a surprisingly large influence on the local
field potential and associated rhythms, such as gamma oscillations, which,
in turn can affect the variability of neuronal spiking (Gómez-Laberge
et al. 2016). Taken together, our findings have led us to focus our
rodent studies on top-down inputs to the layer 1 apical dendrites of
pyramidal cells and to interactions with somatostatin-containing
interneurons.
1)
Feedback and learning. While animals learn to
discriminate the orientation of noisy oriented textures, we record in V1
with multi-electrode arrays to test the predictions of a hierarchical
Bayesian model of perceptual inference. (Ariana Sherdil, Postdoctoral Fellow; Camille Gómez-Laberge, Postdoctoral Fellow; collaboration with Dr. Ralf Haefner and Richard Lange, Univ. of Rochester).
See Lange et al. 2018.
2)
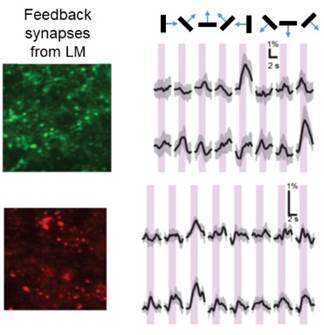
Feedback mechanisms. We have developed a rodent
preparation that lets us simultaneously monitor the activity of both
neuronal cell bodies in V1 and the synapses projecting back to them from
higher visual areas. We do this using genetically encoded Ca++ indicators
of different colors that are conjugated to proteins that traffic them to different
parts of the neuron. This allows us to measure and, ultimately, manipulate
the different sources of information to test circuit-level hypotheses about
how top-down influences affect sensory inputs. (Abhinav Grama, Postdoctoral Fellow; Susanne Haridi, Master’s Student; Peter Kim, Harvard undergraduate)
3)
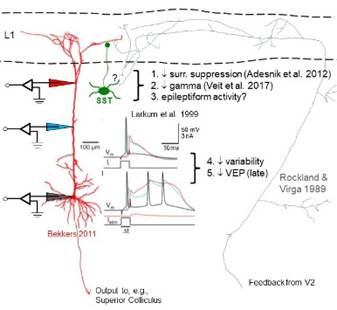
Layer 1 connectome. This project, motivated by
our recent studies of the effects of V2 inactivation on the response
properties of V1 neurons in awake, behaving monkeys, is still in the
planning stages. Stay tuned! (collaboration with Kathy Rockland, Boston University; HMS labs of Gord Fishell, David Ginty and Wei-Chung Allen Lee)
|